В теле рыбы различают голову, туловище и хвост, границы между которыми сильно варьируют у различных рыб. На голове различают парные глаза, парные или реже непарные носовые отверстия, рот и жаберные щели; на туловище обычно имеются: плавники непарные—спинной и подхвостовой и парные—грудные и брюшные, кожистые выросты, укрепленные внутренним твердым скелетом, играющие важную роль при плавании рыбы; заднепроходное отверстие, которым у огромного большинства рыб туловищная часть и заканчивается; внутри туловища находится полость тела, в которой расположены все органы рыбы. Тело рыбы бывает или голое или покрытое костными образованиями и чешуей, имеющими защитную функцию. Различают несколько типов чешуи: более древнюю плакоидную, представленную мелкими пластинками, скрытыми в коже, от которых отходит направленный назад шипик,—это чешуя акул и скатов; ганоидную, или ромбическую, которой были покрыты многие ископаемые рыбы и немногие из ныне живущих,—чешуи современных высших рыб—циклоидную, с гладким краем и ктеноидную с шиповатым. Между всеми типами чешуй имеются генетическая связь и переходы.
Внутреннюю опору тела рыбы составляет с к е л е т, в котором различают: черепную капсулу (заключает головной мозг), осевой скелет, или позвоночник, проходящий по всей длине тела от головы до хвоста, и внутренний скелет плавников. Начавшись с эластического однородного шнура, или хорды, как это мы находим у круглоротых, позвоночник рыб претерпевает такую эволюцию: хорда окружается скоплениями хряща или даже замещается хрящом, обычно распадаясь на отдельные позвонки, пропитанные известью, как это имеет место у хрящевых рыб; у высших рыб позвоночник окостеневает, распадаясь на хорошо скрепленные друг с другом костные, обычно двояковогнутой формы позвонки, между которыми в вогнутых частях сохраняются остатки хорды. От позвонков в передней части тела отходят удлиненной формы костные образования, или ребра, укрепляющие стенки полости тела ж предохраняющие внутренние органы от сдавливания. Каждый позвонок в своей верхней части образует хрящевые или костные дуги, внутри которых проходит спинной мозг; в задней, хвостовой части позвоночника позвонки рыб. образуют нижние дуги, где проходят кровеносные сосуды.
Череп высшей костистой рыбы представляет комплекс многочисленных косточек, тесно соединенных друг с другом. В нем отличают кости обонятельной, глазной, слуховой и затылочной частей головы.
Если сравнить череп высшей костистой рыбы с черепом низших рыб, то в глаза бросятся сильная дифференциация и специализация черепа первой и резкое отличие его не только от черепа круглоротых, но и хрящевых. Если же мы будем изучать развитие черепа костистой рыбы, то - убедимся, что в зародышевой жизни череп каждой костистой рыбы проходит последовательно стадии развития, свойственные низшим хордовым и низшим рыбам: сначала закладывается хорда, не расчлененная на сегменты, и еще нет расширенного головного конца в передней ее части; в следующих стадиях происходит дифференцировка передней части осевого скелета и образование первичного черепа, еще не образующего сплошной коробки, а состоящего из отдельных капсул—обонятельной, зрительной и слуховой. Если первая фаза развития соответствует состоянию внутреннего скелета, наблюдаемого у бесчерепных {ланцетника и его предков), то вторую стадию развития мы можем сопоставить с развитием черепа у круглоротых (миноги, Максины и их предки—первично-черепные); в следующих фазах своего развития череп костистой рыбы можно сравнивать с состоянием черепа у хрящевых рыб; в стадиях личиночной и мальковой происходит уже окончательное формирование внутреннего скелета: сплошной хрящевой внутренний скелет постепенно окостеневает отдельными костями, причем у низших костистых первичный хрящевой скелет (лососевые, сельдевые) в значительной степени сохраняется и у взрослой рыбы, обнаруживаясь при препаровке под поверхностными кожными и хрящевыми костями. Образование вместо массивного хрящевого черепа относительно небольшого и более легкого костного, несомненно, сыграло свою полезную роль в жизни рыбы, улучшив условия плавания рыбы, сделав ее тело более подвижным, ловким, а следовательно, более приспособленным к улавливанию добычи самого разнообразного характера. Распадение сплошного хрящевого черепа на большое число плотно соединенных косточек имело благоприятное влияние и на развитие важнейшего органа рыб—головного мозга, представляя более надежную защиту этому важному органу. Плавники представляют образования,- чрезвычайно приспособленные для плавания: это тонкие легкие пластинки, в большинстве случаев играющие роль направляющих килей; если меняется способ передвижения рыбы, плавники могут до неузнаваемости изменять свою форму, превращаясь в присоски, органы защиты и привлечения добычи; в некоторых случаях на их свободных концах развиваются светящиеся органы (глубоководные рыбы), а у тех рыб, которых мы называем летающими, парные плавники развиваются в огромные своеобразные «парашюты», помогающие рыбе благополучно спланировать на поверхность воды. Во всех этих случаях изменяется не только внешняя форма плавников, но и весь их внутренний скелет, характер расположения и действия мышц, приводящих в движение эти плавники, а также характер их иннервации.
Если сравнить анатомию скелета плавников низших рыб и высших костистых, то и здесь наблюдается тот же процесс дифференциации и специализации, какой мы отметили в черепе и осевом скелете костистых рыб по сравнению с низшими рыбами: плавники высших рыб допускают гораздо более обособленные и разнообразные движения, чем у низших рыб.
Костяк рыбы находится в тесной зависимости от более или менее мощного слоя мышц, расположенного между кожей и скелетом. Разнообразные движения зависят от сократительной деятельности клеток этого слоя, называемого в обиходе «мясом» рыбы, о двигательной функции которого будет сказано ниже. Составляя слой стенок полости тела, мышечный слой играет немалую предохранительную для внутренних органов тела роль, защищая эти органы от сдавливания и сотрясения.
Скелет окуня (Регса fluviatilis).
Лобная кость. 2—Предлобная кость. 4—Заднелобная кость. 5—Нижняя затылочная кость (лежит под костью 28). 6—Клиновидная кость. 7—Теменная кость. 8—Остистый отросток затылочной кости. S—Наружная затылочная кость. 12—Сосцевидная кость. 14—Глазной отросток клиновидной кости (не виден). 15—Передняя клиновидная кость (не видна). 16—Сошник (скрыт под костью 19). 17—Межчелюстная кость. 18—Верхнечелюстная кость. 19—Подглазничная кость. 19 —Задние глазничные кости. 20—Носовая кость. 21—Подвисочная косточка. 22—Небная кость (перед 24, скрыта под 19). 23—Височная кость. 24—Поперечная кость. 25—Внутренняя крыловидная кость. 26—Скуловая кость 27.—Барабанная кость. 28—Жаберная крышка. 30—Преджаберная кость. 31—Соединительная кость. 32—Поджаберная кость. 33—Межжаберная кость. 34—Зубная часть нижней челюсти. 35—Суставная часть нижней челюсти. 36—Угловая часть нижней челюсти.
Позвонки—а, а, а. Сросшиеся хвостовые позвонки—Ь. Поперечные отростки, большею частью отделенные от тела.—с,с. Сросшиеся поперечные отростки под хвостом, между которыми остается отверстие для аорты,—(1,(1. Реберные придатки—Г,I. Остистые отростки—g,g. Передние вставочные косточки—h,h. Задние вставочные косточки—i,i. Твердые лучи первого спинного плавника (числом 14)—к,к. Твердые лучи (числом 3) второго спинного плавника—г,г. Мягкие лучи (числом 12) второго спинного плавника—m,m. Короткие твердые лучи хвостового плавника—п,п. Мягкие лучи хвостового плавника—о. Нижние остистые отростки—р,р.
Нижние вставочные косточки—g.g (у заднепроходного плавника. Твердые лучи заднепроходного плавника—г. Мягкие лучи заднепроходного плавника—я. Лопатка, раздающаяся на две части—АВ. Плечевая кость—С. Плечевой пояс—ABC. Локтевая кость—D. Лучевая кость—В. Четыре косточки запястья—F. Луч плавника, сочлененный е лучевой костью—о. Разветвленные лучи плавника—Н. Две косточки, называемые клювовидным отростком и ключицей,—I К. Задняя конечность—L, Твердый луч брюшного плавника—N. Y брюшного плавника—N. Y брюшного плавника—М. Мягкие лучи.
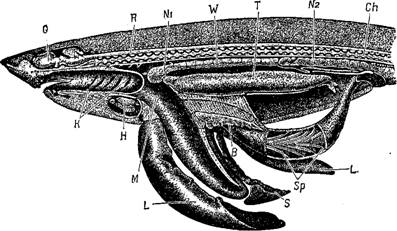
Разрез тела акулы-самца. В—брюшная железа; Ch—позвоночник с хордой; G—головной мозг; Н—сердце; К—жаберные щели; Ъ— печень; М—желудок; N,—семепроводящая часть почки; N,—-выделительная часть почки; R—спинной мозг; S—селезенка; Sp—спиральный клапан; Т—семенник; W—семепроток.
Внутренние органы рыбы соединяются в ряд систем, тесно связанных между собой. От согласованной и непрерывной деятельности их зависит правильность работы всего организма. Остановимся вкратце на главнейших из них.
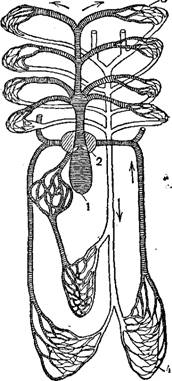
Кровеносная система рыб представлена: 1) двухкамерным сердцем, состоящим из мускулистого желудочка и тонкостенного легко спадающегося предсердия; только у двоякодышащих рыб, представляющих уже переход к амфибиям, начинает образовываться третья камера,— в предсердий появляется неполная перегородка, которая при сокращении предсердия разделяет его на правую и левую половины; 2) артериальными кровеносными сосудами с толстыми неспадающимися эластическими стенками (кровь перемещается от сердца); 3) венозными сосудами с тонкими легко спадающимися стенками (кровь идет к сердцу); 4) венозным расширением (пазухи), в которое впадает кровь, прежде чем войти в предсердие; венозная пазуха имеет тоже тонкие спадающиеся стенки.
Кровеносная система рыб является замкнутой, т. е. артериальные сосуды, принося кровь в тот или другой орган, распадаются на сеть микроскопически малых сосудов, или капилляров, имеющих стенку из одного ряда клеток, через которую легко диффундируют газы и жидкости.
Артериальные капилляры, отдав органу кислород, питательные вещества и получив от него продукты распада и углекислоту, непосредственно переходят в венозные капилляры, которые сливаются в венозные сосуды. По мере движения к сердцу последние все укрупняются от вливающихся все новых вен и впадают в венозную пазуху или в предсердие двумя парами крупных венозных сосудов: парой верхних кардинальных вен, несущих кровь от головы и передней части тела, и парой нижних кардинальных, несущих венозную кровь из задней части туловища и от внутренностей.
Верхние и нижние кардинальные вены, сливаясь вместе, образуют парные Кювьеровы протоки, по которым венозная кровь притекает в венозную пазуху и оттуда в предсердие.
Только у двоякодышащих мы встречаемся с образованием нижней полой вены, свойственной уже всем вышестоящим позвоночным.
Кровь в сердце рыб поступает венозная (исключая двоякодышащих, где благодаря легочному дыханию она смешанная). Из сердца кровь гонится через желудочек в жабры, где приносящие сосуды распадаются на густую сеть капилляров и где происходит газообмен с омывающей жабры водой; из воды поступает кислород, а отдается углекислота. Из жабер кровь, уже богатая кислородом, поступает в спинную аорту и разносится по всему телу. Непрерывность движения крови в теле рыбы обусловливается ритмическим сокращением желудочка сердца и всасывающей деятельностью сердца благодаря периодическому спадению и расширению венозных сосудов, венозной пазухи и предсердия.
Кровь у рыб красного цвета и заключает в себе: 1) красные кровяные тельца, переносящие кислород, поглощенный кровью; 2) белые кровяные тельца, играющие защитную роль против проникновения в кровь всяких вредных начал; 3) плазму, заключающую в себе все необходимые для жизни рыбы питательные и пластические вещества. В кровь же поступают и жидкие и газообразные продукты обмена веществ различных тканей. Кровь, таким образом, представляет внутреннюю, так сказать, среду всего организма, без которой не может протекать обмен веществ.
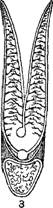
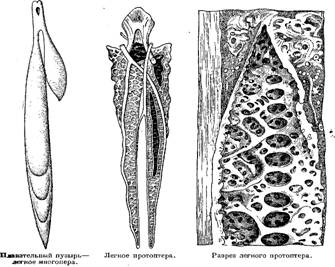
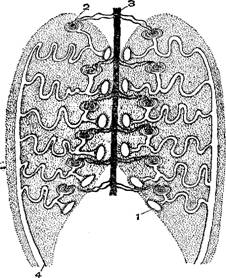
Органы дыхания рыб представлены жабрами, имеющими разную форму—складчатых внутри мешочков, пластинок, лепестков, пучков и т. п., пронизанных густейшей сетью мельчайших кровеносных сосудов, входящих в непосредственное соприкосновение с водой, омывающей жаберную полость. По своему устройству жабры представляют огромную поверхность соприкосновения с водой, приносящей необходимый для дыхания кислород, поглощаемый непрерывно притекающей к жабрам кровью и разносимый по всему телу. У двоякодышащих и кистеперых рыб к жаберному дыханию присоединяется и воздушное при посредстве плавательного пузыря, превратившегося в своего рода легкое.
Органы пищеварения рыб представлены: 1) пищеварительным трактом, 2) печенью, 3) поджелудочной железой и 4) рядом лимфатических желез, способствующих своими секреторными выделениями дальнейшему изменению переваренной пищи.
В пищеварительном тракте различают: ротовую полость, обычно вооруженную зубами, глотку, прорезанную с боков жаберными щелями, пищевод, желудок, где у большинства рыб начинается пищеварительный процесс, кишечник, где продолжается процесс переваривания пищи и ее всасывание. Заканчивается пищеварительный тракт заднепроходным отверстием и только у двоякодышащих и хрящевых—клоакой. Процесс переваривания включает в себе два противоположных процесса:
процесс изменения белков, жиров и углеводов пищи и приведение их в растворимое состояние и процесс созидания в стенках кишечника и в щетках тканей из этих измененных частей пищи свойственных данной рыбе белков, жиров, углеводов и других соединений. Все эти изменения элементов пищи происходят пол влиянием так называемых ферментов:
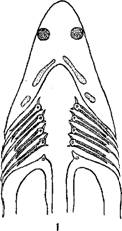
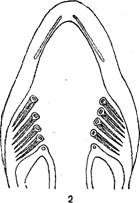
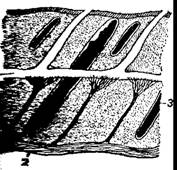
Жабры рыб.
1—акулы; 1—костистой рыбы; 3—жаберный лепесток.
пепсина, выделяемого клетками желудка, разлагающего в кислой среде белки пищи до пептонов, трипсина, выделяемого поджелудочной железой, влияющего на разложение белков, стеапсина, способствующего перевариванию жиров, и, вероятно, ряда других ферментов. Надо полагать, что и синтезы измененных частей пищи в новые свойственные данной рыбе соединения происходят не под влиянием специфических ферментов.
Переваренная и соответственным образом измененная пища поступает в лимфатическую и кровеносную системы и разносится по всему телу рыбы, принося каждому органу, каждой клетке необходимые для нее вещества.
В устройстве органов пищеварения у рыб мы находим большое разнообразие, в значительной мере обусловленное приспособлением пищеварительного тракта к определенному характеру питания. Известно, что рыбы, питающиеся по преимуществу растительной пищей, имеют более длинный пищеварительный тракт, чем хищники, питающиеся рыбной и другой животной пищей. Но, помимо бросающегося в глаза удлинения и укорочения пищеварительного тракта приспособительно к характеру пищи, в нем происходил, несомненно, ряд глубоких приспособительных изменений в микроскопическом устройстве стенок пищеварительного тракта, в выработке его клетками специфических для данной пищи соединений.
Пищеварительный тракт рыб приспособлен к освоению организованной пищи из окружающей водной среды. Усвоение растворов азотистых соединений, вероятно, также имеет место: вместе со всасыванием неорганических солей, вероятно, всасываются и эти соединения, но сколько-нибудь существенного значения они не имеют, ибо обычно содержатся в воде в ничтожных количествах, и для организма рыбы гораздо выгоднее было приспособиться к использованию концентрированных в организмах пищевых элементов, чем производить огромную работу по вылавливанию и концентрации их из растворов.
Непосредственный анализ пищи рыб ясно показывает, что как растительные, так и животные организмы составляют основную пищу рыбы и с жизнью этих организмов теснейшим образом связывается и жизнь самой рыбы. Мало того, как показывает опыт, каждая рыба приспосабливается к определенной пище, и из обычно большого количества организмов, которые встречаются в пищеварительном тракте ее, можно выделить такие организмы, которые встречаются в преобладающем количестве и при наличии которых рыба наилучше бывает упитана. Эти организмы составляют основную, или главную, пищу рыбы, к которой приспособлены ее пищеварительный тракт и весь ее организм. При отсутствии главной пищи рыба может существовать и за счет организмов, обычно играющих второстепенную роль в питании ее, но тогда она не бывает так упитана, как при «обычной» пище, и не так быстро растет.
Такую пищу не совсем правильно зовут «случайной», так как вряд ли случайным может считаться захватывание ее рыбой вместе с главной пищей: вернее, эта второстепенная пища заключает в себе некоторые добавочные вещества, необходимые для питания рыбы.
Иногда рыба, за отсутствием свойственной ей пищи, вынуждена питаться необычной для нее пищей. Эта вынужденная пища, правда, может поддерживать существование рыбы, но не дает прироста и упитанности телу рыбы.
Эффективность питания разными родами пищи обусловливается не только наличием или недостатком в ней тех или других питательных веществ: белков, жиров и углеводов, но и приспособленностью или неприспособленностью самого пищеварительного тракта рыбы к той или другой пище. Иногда, вероятно в связи с состоянием самого организма рыбы, меняется и ее пища.
Выделительные органы, или почки, рыб. Поступающие в кровь продукты обмена, мочевая кислота, мочевина и пр., выделяются из крови при посредстве почек. Почки, расположенные с той и другой стороны позвоночника, представляют парные органы рыхлой консистенции, приобретающие кровяной цвет от огромного количества протекающей через них крови.
Следует отметить, что каждый входящий в почки сосуд распадается на двойную сеть капилляров, благодаря чему создаются наилучшие условия для фильтрации. Образуется так называемая воротная система кровообращения.
При ознакомлении с микроскопическим устройством почек у рыб мы замечаем в них, помимо свойственной почкам специфической соединительной ткани, огромное количество капилляров, составляющих главную массу вещества почек. В теснейшем контакте с капиллярами находится система многочисленных выделительных, или мочевых, канальцев. В своем полном виде мочевой каналец состоит из нескольких частей: начинается он расширенной частью—реснитчатой воронкой, благодаря движению ресничек которой жидкость всасывается в каналец; несколько отступя от реснитчатой воронки, мочевой каналец образует новое расширение—бауманову капсулу, имеющую вид шара со вдавленными внутрь стенками; в эту капсулу и входят вены и артерии и распадаются там на капилляры и отфильтровывают жидкие продукты распада в отходящий от капсулы мочевой каналец. Отфильтрованная жидкость движением ресничек клеток, выстилающих стенки мочевого канальца, гонится к мочеточнику, в который впадают, в конце концов, все выделительные, или мочевые, канальцы. По мочеточнику выделенная жидкость выводится наружу или через клоаку у низших рыб, или через отверстие на мочеполовом сосочке. Фильтрация происходит ритмически и беспрепятственно благодаря тому, что кровь в капиллярах протекает под достаточным давлением, а сама отфильтровываемая жидкость представляет собой соли кристаллоидов, т. е. таких веществ, которые, в противоположность коллоидам, легко профильтровываются через стенки капилляров. Клетки, выстилающие стенки мочевых канальцев, играют активную роль, высасывая из фильтра обратно в капилляры необходимые для крови соли, которые механически отфильтровались раньше и, таким образом, способствуют восстановлению нормального Мальпигиево тельце, солевого состава крови, что, как известно, чрезвычайно 1—клубочек кровеносных сосудов; 2—бауманова капсула важна для всех процессов обмена в организме рыбы: з—каналец.
ведь от количества солей в крови в прямой зависимости стоит и величина осмотического давления, без которого, как известно, невозможна ни жизнь отдельных клеток, ни целого организма рыбы.
Таким образом, почки у рыб являются не только органами выделения, но и регуляторами солевого состава, а вместе с тем и осмотического давления крови. Та тесная связь между мочевыми канальцами и капиллярами, которую мы встречаем в почках рыбы, говорит о постоянном взаимодействии между фильтратом мочевых канальцев и крови, протекающей по капиллярам.
Почки рыб расположены в туловищной части тела и называются туловищными или вторичным, и в отличие от головных, или первичных, свойственных бесчерепным (ланцетнику), всем рыбам в эмбриональный период их жизни и только немногим рыбам во взрослом состоянии.
Третичных, или тазовых, почек, свойственных всем высшим позвоночным, у рыб еще не появляется.
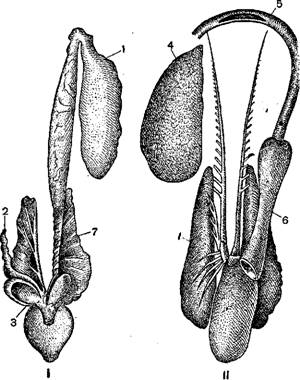
Половые органы рыб расположены в полости тела и находятся обычно в тесной связи с почками, канальцы которых частью служат для выведения наружу половых продуктов. У большинства рыб половые органы парные. Женские называются яичниками и содержат в себе более или менее многочисленные яйца или икринки; мужские называются семенниками, или молоками, и содержат огромное количество семенных нитей, или сперматозоидов. В зависимости от продолжительности цикла жизни половые продукты рыб созревают не в одинаковое время: есть рыбы, которые делаются половозрелыми в возрасте менее года (некоторые мелкие бычки), но зато имеются и такие, у которых половая зрелость наступает не ранее 15 или 16 лет (белуги и калуги). Только зрелая икра может быть оплодотворена и дать начало новому организму.
Период размножения происходит обычно раз в году. Однако некоторые тропические рыбки при благоприятных условиях могут нерестовать несколько раз в году, так как новые генерации икры созревают очень быстро у таких рыбок.
Размножение является одним из важнейших актов, которым поддерживается существование вида и особи. Естественно, что у рыб, для сохранения развивающейся икры и вышедшей из нее молоди, выработался целый ряд полезных приспособлений. У большинства рыб икра откладывается наружу и поливается там молоками самца; только у немногих происходит внутреннее оплодотворение икринок, и развитие протекает внутри тела самки; молодые рыбки при этом выводятся через половое отверстие наружу. У одних рыб икра откладывается на дно, у других свободно плавает; некоторые рыбы прикрепляют оплодотворенную икру к растениям на дне, зарывают в песок или даже устраивают для нее особое гнездо, которое и охраняется родителями нередко в течение всего развития молоди (колюшки), некоторые охраняют отложенную икру (маслюк). У тропического сома (аспредо) икра прикрепляется после оплодотворения на брюшке самки, у которой ко времени нереста брюшко приобретает особый губчатый характер, благодаря чему икринки при надавливании на них брюшка рыбы легко вдавливаются внутрь губчатой ткани; но приспособление рыбы к вынашиванию икры этим не ограничивается: к каждой икринке прирастает особый стебелек, богатый кровеносными капиллярами, очевидно, приносящий развивающемуся в икринке зародышу все необходимое для жизни.
Не менее удивительное приспособление для развития оплодотворенной икры мы встречаем у самцов морского конька и морской иглы, у которых в период нереста развиваются на брюшке кожистые складки, в которые самками откладываются оплодотворенные икринки; стенки этих складок делаются губчатыми и богатыми кровеносными сосудами. У самца морского конька они замыкаются в мешок, так называемый «зародышевый», или «зародышевую камеру»; у морских игл замыкания стенок складок не происходит, но в том и другом случае развивается в стенках этих складок губчатая, обильная кровью ткань; икра получает не только безопасное убежище на все время развития, но и все необходимое для нормального развития. По-видимому, мы можем находить некоторую аналогию между условиями развития икры в таких замкнутых камерах, как зародышевая камера морского конька, и условиями развития икры у живородящих рыб внутри полости тела.
Не менее любопытны приспособления у некоторых рыб (хилодиптерус, паратилапия и другие) для вынашивания оплодотворенной икры в ротовой полости; при этом в ротовой полости рыбы образуется особое мешкообразное углубление, в котором благодаря постоянному току воды икринки находятся в непрерывном вращательном движении до выхода из них мальков, что чрезвычайно полезно для развивающегося зародыша в отношении равномерного снабжения икринок свежей водой. Имеются указания, что и самец принимает участие в вынашивании икры, периодически сменяя утомившуюся самку и производя те же манипуляции с икрой, что и она. Вышедшие из икры мальки первое время тоже охраняются родителями, которые при наступлении опасности втягивают их с водой в ротовую полость. Вынашивание икры на теле родительских форм является актом, благоприятствующим сохранению развивающейся икры; еще более целесообразным актом является живорождение, при котором икра оплодотворяется внутри тела самки. Живорождение широко распространено у хрящевых рыб (акул и скатов), у которых выработался целый ряд приспособлений для оплодотворения и развития икры внутри тела матери: наружные совокупительные органы самцов, «матка», сперматофоры и пр. Живорождение очень часто встречается и у представителей семейства зубатых карпов, где в некоторых случаях благодаря ему сохраняется существование вида. Среди других костистых рыб имеются и живородящие формы (циматогастер, бельдюга, норвежский окунь и другие скорпеновые).
Можно привести еще немало примеров приспособлений, полезных для успешного развития оплодотворенной икры, но мы закончим замечательным способом отложения, оплодотворения и развития икры у одной маленькой, но широко распространенной в водах СССР рыбки—горчака. Горчак использует для развития своей икры двустворчатых моллюсков Unio и Anodonta, в полость мантии которых самка откладывает зрелую, но еще неоплодотворенную икру при помощи особого яйцеклада, развивающегося у рыбы к моменту созревания икры. После отложения икры самкой в тело моллюска самец горчака, присутствующий при этом, немедленно выпускает в воду около моллюска.
порцию молок, которые втягиваются вместе с водой через водный сифон в полость мантии моллюска и оплодотворяют отложенную туда самкой икру. Развитие икры происходит в чрезвычайно благоприятных условиях при постоянном, ритмическом притоке свежей воды, втягиваемой регулярно моллюском. Как исторически вырабатывалось это приспособление, связавшее так тесно существование горчака с существованием названных выше моллюсков, пока остается неизвестным.
Всех рыб, имеющих те или другие полезные для развития икры и мальков приспособления, не совсем правильно называют «имеющими заботу о потомстве», противопоставляя их таким, которые этой «заботы» не обнаруживают. Но закономерность в отношении количества откладываемой икры между рыбами этих двух категорий действительно существует: там, где выработались те или другие полезные приспособления для развивающейся икры и мальков, будет ли это устройство гнезд, прикрепление икры к телу родителей или живорождение, количество икринок незначительно и исчисляется десятками, сотнями или немногими тысячами; у тех же рыб, у которых оплодотворенная икра просто откладывается в водную среду, обычно количество икринок насчитывается сотнями тысяч и даже многими миллионами (тресковые, луна-рыба, камбаловые и другие).
Если из икры выходит молодая форма, мало отличающаяся от родительской или быстро приобретающая все основные признаки ее, то говорят, что данная рыба имеет прямое развитие. Если из икры выходит молодь, отличная по внешним признакам от взрослой родительской формы, то тогда говорят, что данная рыба имеет личиночную стадию развития, обычно продолжающуюся значительный промежуток времени (у угря 3 года, у миног не менее 2—3 лет). В течение этого времени в организме личинки происходят коренные, глубокие изменения, после чего она приобретает все признаки родительской формы.
В некоторых случаях изучение личиночных стадий развития позволяет судить о филогении данной рыбы (личиночные стадии морского черта, луны- рыбы и др.).
Наступление половозрелости у рыб зависит в значительной степени от цикла жизни их: при коротком цикле рыба делается способной к размножению на первом-втором году (хамса, килька и другие), обычно на третьем, четвертом, пятом (карповые, некоторые лососевые, сельдевые); при продолжительном цикле жизни, как у осетровых, нельмы и некоторых других морских рыб, половозрелость наступает много позже—в возрасте от 10 до 16 лет.
Впрочем, сроки наступления половозрелости даже для одного и того же вида в различных районах его распространения неодинаковы. Первые годы половозрелая рыба обычно нерестует ежегодно, но затем к старости промежуток между двумя соседними нерестами увеличивается. Только немногие рыбы—дальневосточный лосось, некоторые из южных сельдей («залом», или черноспинка), миноговые—нерестуют раз в жизни и после первого нереста погибают.
Нервная система рыб. Центральным органом, управляющим работой всех органов рыбы и регулирующим эту работу, является нервная система, представленная головным, спинным мозгом и симпатической нервной системой, расположенной по бокам позвоночника и автоматически регулирующей деятельность внутренних органов.
Основным элементом нервной системы является Нервная клетка со всеми ее отростками (так называемый «нейрон»), воспринимающая раздражения извне и от внутренних органов и передающая в ответ на эти раздражения импульс .мышцам и органам тела.
Различают нервные клетки, воспринимающие раздражения, не совсем правильно называемые «чувствующими», и клетки, передающие раздражения мышцам, железам, коже и внутренним органам, называемые «двигательными».
Между «чувствующими» и «двигательными» клетками бывает или постоянное соединение или только временный контакт, благодаря этому одна и та же «чувствующая» клетка может войти в контакт в самых разнообразных комбинациях с многочисленными «двигательными» клетками, так что раздражение, полученное извне одной клеткой, может по мере надобности передаваться и мышцам и целому ряду внутренних органов. Нервные клетки в мозгу рыб насчитываются многими миллионами; если вспомнить, что каждая клетка это центральная станция, принимающая внешние и внутренние раздражения и вызывающая целесообразные ответные реакции организма в форме того или иного движения, то нам станет понятным все значение этих «бесчисленных нервных станций» в борьбе за существование и в выживании вида (сама нервная система также развилась путем отбора в борьбе за существование).
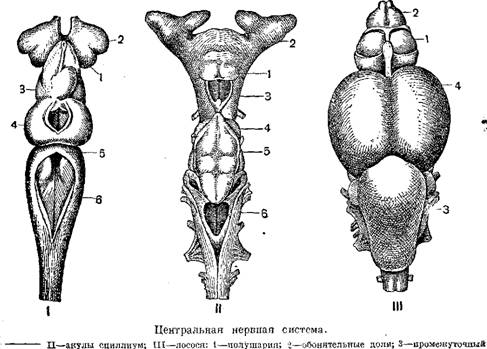
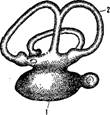
В связи с невысокой степенью развития организации рыб их головной мозг сохраняет еще примитивные черты, свойственные низшим позвоночным. Он имеет линейное расположение частей (передний, промежуточный, средний и задний мозг), не имеет еще объединяющего все части головного мозга центра, каковым у высших позвоночных являются полушария переднего мозга.
Кора больших полушарий головного мозга у рыб вместо нервной ткани имеет еще эпителиальный слой («паллиум»); только у двоякодышащих кора больших полушарий имеет нервные клетки, частично нервная ткань встречается и в коре полушарий акуловых.
По величине головной мозг рыб очень мал и никогда не заполняет целиком полости черепной коробки. По сравнению с мозгом наземных позвоночных головной мозг рыб составляет относительно малую часть веса тела; так, мозг налима составляет всего щуки 1/1300, а тунца 37 000 веса тела.
Органы чувств рыбы. Многообразные внешние раздражения воспринимаются так называемыми органами чувств (анализаторами), рецепторами специфических раздражений, тесно связанными с определенными частями головного мозга.
Одни из этих органов воспринимают разного рода механические раздражения, другие воспринимают химические раздражения (органы обоняния и вкуса). И те и другие имеют черты приспособления к восприятию соответствующих раздражений в водной среде, обычно локализуясь на определенных частях тела рыбы. Органами восприятия механического раздражения являются органы зрения, равновесия и слуха, кожные и органы боковой линии. Беглое описание всех этих органов дается ниже.
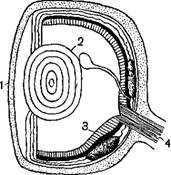
Орган зрения рыб представлен парными глазами, имеющими все те части, которые нам известны и у наземных животных, но приспособленными для зрения в водной среде.
Снаружи глазного яблока расположена плотная оболочка-склеротика (или белковая, у глубоководных рыб, где глаз выдерживает большое давление), сильно укрепленная хрящевыми и костными образованиями, переходящая спереди в прозрачную, плоскую у рыб роговицу, за белковой в последовательном порядке располагаются оболочки: серебристая, сосудистая, пигментная и самая внутренняя— сетчатая. Последняя заключает в себе светочувствительные клетки (палочки и колбочки), многочисленные окончания зрительного нерва, нервные и опорные клетки.
Хрусталик у рыб шарообразной формы, не изменяет своей выпуклости; аккомодация получается благодаря отходящему от сосудистой оболочки серповидному отростку, приближающему хрусталик к сетчатой оболочке. Орган зрения у рыб в зависимости от их образа жизни подвергается значительным изменениям: у рыб, живущих при полном отсутствии света, он атрофируется и даже совсем исчезает; у рыб, которые живут в среде с недостаточным светом, наоборот, глаза увеличиваются или даже делаются (у некоторых морских глубоководных рыб) телескопическими, благодаря чему могут быть использованы даже немногочисленные световые лучи. К замечательным приспособлениям рыбы к использованию световых лучей в недостаточно освещаемой среде следует отнести появление особого светящегося органа внутри глазного яблока, поддерживающего сетчатку в состоянии постоянного возбуждения и делающего ее более восприимчивой к восприятию даже очень слабых лучей извне; такое внутреннее освещение сетчатки известно у некоторых глубоководных рыб. Нервные центры, которым передаются из глаза световые раздражения, находятся в среднем мозгу.
Орган обоняния, играющий в жизни рыбы большую роль, представлен парными (реже одиночными) ямками в передней части головы, слизистая оболочка которых образует многочисленные складки (увеличение обонятельной поверхности), богатые обонятельными клетками. К этим клеткам подходят ветви обонятельного нерва и оплетают их густой сетью своих телодендриев; обонятельные раздражения передаются по ветвям обонятельного нерва центральным клеткам, находящимся в передней Доле головного мозга. У тех рыб, у которых обоняние играет важную роль в отыскании добычи,—акуловые, миноговые, миксиновые и некоторые другие,—обонятельные доли головного мозга сильно развиты, богаты воспринимающими обонятельные раздражения нервными клетками, вступающими по мере надобности в контакт с разнообразными двигательными центрами.
Органы вкуса представлены у рыб так называемыми вкусовыми почками, находящимися в слизистой оболочке рта, глотки, жаберных дуг, губ и в коже покровов всего тела. Почки эти имеют вид овальных углублений, выстланных чувствительными и опорными клетками, оплетаются окончаниями языко-глоточного, блуждающего и лицевого нервов, устанавливающих связь с задними частями головного мозга (продолговатый мозг и мозжечок), где и находятся центры вкуса.
Органы равновесия и слуха представлены в виде одного внутреннего уха высших позвоночных: ни наружного уха, ни улитки у рыб нет. Орган слуха расположен в полостях слуховых костей и представляет кожистый орган, состоящий из большого нижнего и овального верхнего мешочков, от верхнего отходят три полукружных канала, от нижнего—небольшой отросток: зачаток будущей улитки. Пространство между костными стенками и перепончатым органом заполнено наружной лимфой, а внутри органа внутренней лимфой, стенки последнего выстланы чувствительным эпителием, в связь с которым и вступают нервные окончания слухового нерва. Всякое нарушение равновесия рыбы вызывает приток внутренней лимфы к определенным частям слухового аппарата, раздражение чувствительных клеток и стремление рыбы восстановить нормальное положение.
Есть основание предполагать, что и чисто звуковые волны до некоторой степени доступны органу слуха рыб. Центры слухового органа находятся у рыб в продолговатом мозгу.
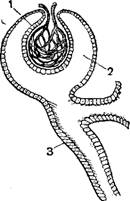
1- линии рыбы. 2—нерв; 3—чешуи.
Органы бокового чувства представляют своеобразный анализатор рыб, неизвестный у наземных животных, за исключением древнейших амфибий и современных в личиночной стадии. Эта так называемая боковая линия представляет собой канал, идущий по бокам тела и головы рыбы в коже и открывающийся наружу рядом отверстий; внутри этот канал выстлан клетками чувствительного эпителия, к которым подходят окончания веточек блуждающего или лицевого нервов. Боковая линия имеет чрезвычайно важное значение для жизни рыбы: благодаря ей, рыба различает токи воды малой напряженности и может ориентироваться в пространстве; даже будучи ослеплена, рыба находит путь из моря в нерестовые реки и т. п.
Органом движения рыб служит мышечная система. Различают мышцы так называемых непроизвольных движений внутренностей (волокна их гладкие) и мышцы поперечнополосатые—мышцы туловища и головы, а также мышцы сердца. Каждая мышца состоит из большого числа сократимых которых и составляет работу мышцы; мышца может проявить тем большую силу и продолжительность работы, чем большее количество волокон входит в ее состав и чем короче эти волокна. Если оценивать рыбу как движущийся механизм, то она устроена с точки зрения механики движения выгодно: основную массу ее тела составляют два мощных боковых мускула, тянущихся от головы до хвоста и поделенных соединительнотканными прослойками на ряд сегментов, что весьма выгодно, ибо действует совокупность сегментов с короткими мышечными волокнами, а не сплошные длинные волокна: это дает возможность рыбе при относительно малых размерах быстро и продолжительно плавать.
Мышечная ткань играет важную роль в жизнедеятельности рыбы: непрерывная работа сердца, все движения, связанные с открыванием и закрыванием рта, заглатыванием воды, проталкиванием ее в жаберную полость и выбрасыванием наружу, выделительная деятельность желез, все движения пищеварительного аппарата, имеющие столь важное значение для усвоения пищи (перистальтика и антиперистальтика кишечного тракта), все перемещения в пространстве, которыми обусловливается благосостояние рыбы, а часто и самое ее существование,—все это зависит от работы мышц. Мышечная ткань теснейшим образом связана с нервной системой, составляя в рефлекторном аппарате его конечную часть. И эмбриологически мышечное волокно связано с нервным с самого начала своего возникновения. Мышцы образуют самую мощную часть тела рыбы, так называемое «мясо», содержащее у некоторых рыб значительные скопления жира, богатого витаминами; кроме того, мышечные пучки находятся в отдельных железах, тканях и органах,—всюду, где сократимость мышц имеет значение для правильной работы органа. Словом, мышцы, входя составной частью в различные органы рыбы, являются совершенно необходимыми для жизни всего организма.
Органы внутренней секреции у рыб. Как и у других позвоночных, у рыб имеется ряд желез, лишенных выводных протоков. Вырабатываемые ими вещества поступают непосредственно в кровь и оказывают влияние на весь организм или на отдельные органы рыбы. Выделение таких желез называется гормонами, или инкретами, а самый процесс выделения— внутренней секрецией. Любопытно отметить, что гормоны какой-либо железы на одни органы действуют возбуждающим, на другие угнетающим образом.
Благодаря поступлению в кровь гормонов между различными органами,, помимо нервной, устанавливается еще и химическая, гормональная связь.
У рыб, как и у высших позвоночных, и железы с протоками, имеющие в работе организма свою определенную функцию, несут наряду с этой главной функцией еще и побочную —гормональную. Так, половые железы выделяют мужской и женский половые гормоны, которыми обусловливаются вторичнополовые признаки и брачный наряд рыб. И этим гормонам некоторые исследователи приписывают импульсы к нерестовым миграциям рыб, половые продукты которых еще не достигли полного созревания.
Печень, играющая столь важную роль в процессах пищеварения выработкой желчи, является и железой внутренней секреции. Внутрисекреторная деятельность печени заключается в превращении гликогена, отложенного кровью в ней, обратно в виноградный сахар и выделении его по мере надобности в кровь.
Поджелудочная железа, дающая важнейшие пищеварительные ферменты, как железа внутренней секреции, выделяет гормоны, мешающие почкам выделять из крови сахар, а также способствующие «гликогенной» деятельности печени.
Все железы внутренней секреции, известные у высших позвоночных, имеются в более или менее развитом состоянии и у рыб, являясь, конечно, нетождественными по составу, но одинаковыми по функции. Введение рыбам гормонов от высших позвоночных имеет те же последствия, как и у этих последних; так, гормон полового созревания—пролан (выделяемый гипофизом), полученный из мочи беременной женщины, будучи-впрыснут рыбе, вызывает созревание ее половых продуктов (как семенников, так и яичников). Об этом ясно говорят опыты, произведённые над рыбами проф. С. Н. Скадовским и его учениками. Этот же гормон, впрыснутый крысе, вызывает тоже ускорение созревания ее половых продуктов.
У рыб известны, хотя и недостаточно изучены, нижеследующие железы внутренней секреции, в собственном смысле этого слова.
Щитовидная железа представляет небольшую железу без протока, вырабатывающую гормон—тироксин и, вероятно, еще другие гормоны, имеющие отношение к обмену веществ. Железа эта у всех позвоночных, где она более или менее развита, чрезвычайно обильно снабжается кровью. Удаление щитовидной железы вызывает у всех позвоночных расстройство обмена веществ и гибель оперированного. Тироксин и другие выделения щитовидной железы, вероятно, обязаны своей активностью и тому, что в их состав входит в значительном количестве иод.
Зобная железа присутствует обычно у всех рыб если не во взрослом, то, во всяком случае, в зародышевом их состоянии. Гормоны этой железы имеют влияние на рост молодого организма, на обмен у него солей (кальция и железа) и на созревание половых органов. Расположена парная зобная железа рыб в жаберной полости. Как и у других позвоночных, наибольшего развития она достигает в молодом возрасте, уменьшаясь, а иногда совсем исчезая с наступлением половозрелости.
Подмозговая железа, или гипофиз, расположена в промежуточном мозге у воронки. Ее гормоны оказывают свое влияние на рост костей: при чрезмерном развитии функции этой железы происходит гигантский рост организма, при замедлении—карликовый рост его. Любопытно отметить, что гигантские рыбы (некоторые акулы, белуги, тунцы и пр.) имеют значительно развитый гипофиз, у мелких рыб (некоторые бычки, тюльки) он относительно мал. Гипофиз имеет влияние и на развитие половых продуктов, вырабатывая особый гормон (пролан).
Надмозговая железа, или эпифиз, имеется у всех рыб;, гормоны ее имеют задерживающее влияние на развитие половых продуктов, развитие вторичнополовых продуктов и пр.
Надпочечные органы у рыб имеются, но слабо развиты, и функция их недостаточно изучена. По-видимому, как и у высших позвоночных, эти органы вырабатывают гормон—адреналин, оказывающий свое влияние на симпатическую нервную систему, гладкую мускулатуру и сужение кровеносных сосудов и играющий, таким образом, огромную роль в регулировании кровообращения. Адреналин влияет также на процесс расщепления гликогена в печени. Кроме приведенных выше желез внутренней секреции, несомненно, у рыб существует ряд других с подобной же функцией, как селезенка и многочисленные лимфатические железы, расположенные в пищеварительном тракте, по-пути движения лимфы, но, к сожалению, пока исследований в этом направлении еще нет, и в отношении рыб о многом приходится пока судить по аналогии е высшими позвоночными.
Несомненно, как и у высших позвоночных, у рыб существует тесное взаимодействие между различными железами внутренней секреции, благодаря чему возможна саморегуляция чрезвычайно важных жизненных процессов путем химического воздействия.
Из всего вышеизложенного явствует, что организм рыбы представляет весьма сложную систему, в которой происходит непрерывная согласованная между собой работа всех ее органов. Подчиняясь определенному ритму работы сердца, зависящему от ряда внутренних и внешних условий, волны крови притекают к каждому органу, к каждой клетке.
Сохранение определенного ритма в работе различных органов обусловливается не только хорошей иннервацией их, но и замечательной приспособленностью их к изменяющимся условиям работы, что, несомненно, казалось результатом длительного влияния естественного отбора на сохранение и усовершенствование полезных функций внутренних органов рыбы.
Ознакомившись с органами рыбы, посмотрим, как реагирует рыба на влияние внешней среды.