Форма тела и движения. Вымершие панцырные земноводные (стегоцефалы) обладали чаще всего саламандрообразной формой тела с короткими массивными конечностями. У современных земноводных можно различить три основные формы, или типа, сложения.
Первый из них—тип тритона с более или менее вытянутым туловищем и хвостом; ноги, если они достаточно развиты, могут поддерживать тело над землей или дном водоема.
Часто наблюдается сильное удлинение тела главным образом за счет длинного хвоста, иногда до угревидной или змеевидной формы; при этом всегда происходит одновременное укорачивание конечностей или только передних, как только задних, или, наконец, обеих в одинаковой степени и даже до полного их исчезновения. Таким образом возникает вторая форма тела—змеевидная, известная у некоторых стегоцефалов, у безногих земноводных и у некоторых хвостатых, как, например, у амфиум; однако у хвостатых земноводных всегда можно различить рудименты конечностей. Такие земноводные могут главным образом плавать, а на суше только ползать на брюхе.
Третий тип сложения—тип лягушек и жаб с плотным, более или менее уплощенным и совершенно бесхвостым телом и хорошо развитыми конечностями, причем задние из них по крайней мере такой же длины, как передние. Крайнее развитие этой формы сложения выражается в сильном удлинении задних ног вместе с соответствующими изменениями в строении таза и развитием мускулатуры. Движения различны в зависимости от степени развития задних конечностей; те земноводные, у которых они лишь немного или вовсе не длиннее передних, способны передвигаться, только ползая, поочередно переставляя ноги; наоборот, чем длиннее задние конечности, тем в большей степени проявляются способности к прыганию.
Все земноводные типа тритонов, как личинки, так и взрослые, а также личинки бесхвостых перемещаются в воде с помощью боковых гребных движений хвоста, имеющего вертикальную кожную кайму. Конечности при плавании этих амфибий играют второстепенную роль и обычно направлены назад и прижаты к бокам тела. Безногие земноводные во взрослом состоянии не живут в воде и если попадают в нее, то движутся с трудом. Амфиумы плавают при помощи змеевидных движений всего тела. Наконец, бесхвостые земноводные двигаются в воде толчками с помощью одновременного сильного разгибания и выбрасывания назад обеих задних ног, снабженных в большей или меньшей степени развитыми плавательными перепонками; и у них при быстрых плавательных движениях передние ноги направлены назад и прижаты к бокам тела.
При медленном передвижении по земле у обыкновенной саламандры передние ноги лишь направляют движение, а задние толкают тело вперед; вытянутое тело животного при этом несколько изгибается змеевидно, но не касается земли. Тонкие, вытянутые, длиннохвостые наземные саламандры (Spelerpes, Chioglossa) могут бегать так же быстро, как ящерицы. Способность к передвижению прыжками широко развита среди многих представителей бесхвостых амфибий.
Многие бесхвостые обладают способностью лазать. В связи с этим у них на концах пальцев имеются особые плоские диски, нижняя сторона которых богата железами, выделяющими липкую слизь. Особые гладкие мускулы делают эти диски плоскими. Подобные пластинки для прилипания независимо возникли в самых различных группах бесхвостых, и в некоторых родах имеются все переходы от видов с йросто закругленными концами пальцев к видам, имеющим хорошо развитые диски. Что для прилипания достаточно и слизистых выделений кожных желез на нижней стороне тела, можно ясно видеть на маленьких травяных и водяных лягушках, жерлянках и тритонах, которые взлезают без особых приспособлений для прилипания по стеклянным стенкам аквариума. У некоторых лягушек, живущих на деревьях, как, например, у африканских хватающих лягушек (Chiromantis), 1 или 2 внутренних пальца передних ног могут противопоставляться остальным, благодаря чему эти земноводные при лазании могут охватывать тонкие ветки. У южноамериканских филломедуз (Phyllomedusa) такая «собенность свойственна внутреннему пальцу не только на передних, но и на задних конечностях.
У летучих лягушек (Rhacophorus) с Зондских островов имеются приспособления для планирующего полета. У них пальцы передних и задних ног соединены сильно развитыми перепонками до самых пластинок, служащих для прилипания. Прыгая с дерева, летучая лягушка по косой линии опускается на землю; при этом она держит конечности прижатыми к телу и растопыривает пальцы; одновременно она сильно раздувает легкие, отчего объем тела увеличивается. Все это создает большую поверхность для сопротивления воздушной среде и способствует в известной мере планирующему полету.
Наконец, многие бесхвостые земноводные могут легко зарываться в землю, для чего им служат сильно развитые пяточные бугры с острым режущим краем, покрытые обычно крепким роговым покровом. Лягушки, вооруженные такими пяточными буграми, как, например, водящаяся у нас чесночница, могут зарываться в рыхлой почве часто на глубину более метра; при этом они уходят под землю всегда задом наперед.
Безногие земноводные роются в земле исключительно мордой. Череп у них очень плотного сложения; ротовое отверстие на нижней стороне головы и отодвинуто назад, благодаря чему устраняется попадание земли в рот. У хвостатых земноводных особых,приспособлений для рытья нет.
Размеры тела как вымерших панцирных земноводных, так и современных представителей этого класса весьма различны. Среди триасовых стегоцефалов известны настоящие гиганты с черепом длиной в 1 метр 25 сантиметров (Mastodontsaurue). Крупнейшая из современных амфибий—японская гигантская саламандра—достигает почти 160 сантиметров в длину; амфиума в сирен достигают 1 метра в длину. Но в том же отряде хвостатых земноводных имеются и карликовые формы. Многие виды центральноамериканских горных саламандр, принадлежащие к роду Oedipus, достигают в длину не более 6 сантиметров, а некоторые всего лишь 4 сантиметров. Таким образом, величина крупнейших представителей отряда хвостатых превосходит длину тела самых мелких видов того же отряда в 40 раз. В отряде бесхвостых земноводных наибольшей величины достигает лягушка-голиаф (Rana goliath); длина превосходит 32 сантиметра. В то же время можно насчитать более 10 бесхвостых амфибий, распространенных в Центральной и Южной А в Африке и тропической Азии, многие виды которых не превышают в 2 сантиметров. Таких карликов мы находим среди родов: древолазов (Dei tes), настоящих квакш (Hyla), ложных жаб (Pseudophryne), листолазов (I lobates) и ряда других. Среди безногих земноводных крупнейшая настоящая червяга (Coecilia thompsoni) достигает почти 120 сантиметров в длину, как самая маленькая земляная червяга (Hypogeophis brevis)—всего 2 сантиметров.
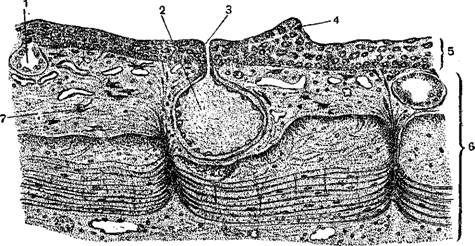
Кожа земноводных может быть названа голой, так как разного роговые образования имеют у современных представителей этого класса незначительное развитие. Обилие слизеотделительных желез обуславливает постоянную влажность и мягкость кожи, что имеет чрезвычайно существенное значение в связи с весьма интенсивным газообменом через кожные покровы дополняющим недостаточно совершенное легочное дыхание наземных Ни одно земноводное не пьет обычным способом; воду земноводные воспринимают исключительно через кожу. Амфибии быстро погибают в условиях недостаточной влажности .от нарушения нормальной деятельности кожных желез, быстрой потери влаги в тканях и нарушения в связи с этим нормального процесса обмена веществ и в первую очередь газообмена.
Кожные покровы земноводных состоят из многослойного плоского эпителия, часто называемого надкожицей, под которой лежит собственно ко или кориум. Последняя образована тремя взаимно перпендикулярными слоями волокон.
У лягушек кожа прикрепляется к телу не на всем своем протяжении а лишь в определенных участках, между которыми располагаются обширные полости, заполненные лимфой.
Ороговение кожи в большей или меньшей степени наблюдается преимущественно у тех амфибий, которые ведут наземный образ жизни. Hanpимер у жаб сильному ороговению подвергаются бородавчатые возвышения спине, концы пальцев, и бугорки на нижней их стороне. Роговые образования, предохраняя нижележащие ткани от механических повреждений и высыхания, в то же время, конечно, затрудняют кожное дыхание, но в таких случаях наблюдается заметно большее развитие легких. Лопатообразные выросты значительных размеров имеются на задних ногах некоторых безхвостых земноводных; они служат, как уже было сказано, для зарывания в землю острыми роговыми когтями на пальцах передних и задних ног обладает уссурийский тритон (Onychodaetylus fischeri). Когти позволяют ему легко удерживаться за подводные предметы на быстром течении. Такие же когти на трех внутренних пальцах задних ног характерны для африканских шпорцевые лягушек (Xenopus). За этими двумя исключениями никакие другие современные земноводные не имеют когтей на пальцах. У некоторых стегоцефалов (в пример Microsauria) имелись настоящие когти.
По мере ороговения и изнашивания внешних слоев эпидермиса происходи его сбрасывание. Линька происходит периодически в течение всей жизни земноводных, но особенно часто у молодых, быстро растущих животных. Надкожица лопается чаще всего на губах или на спине и сбрасывается, как правило, сплошным чехлом, на котором остаются цельными даже чехлики с тонких пальцев.
Окраска кожи амфибий зависит от пигмента, заключенного главным образом в кориуме, но отчасти также и в эпителии. Зернышки пигмента крестного, бурого и черного цветов лежат в особых разветвленных клетках хроматофорах; кроме того, встречаются белые зерна гуанина; желтый пигмеи» диффузно распределяется в роговом слое эпидермиса. Очень подвижные хроматофоры могут не только изменять свою форму, сжиматься, сокращать и вытягивать свои отростки, но в известной мере также перемещаться между клетками кориума и даже проникать в межклеточные пространства эпителия.
Пигментные зерна, в свою очередь, могут изменять свое положение в хроматофорах, то распределяясь в них равномерно, то образуя скопления в разных их частях. Если принять во внимание, что хроматофоры, заключающие пигментные зерна разных цветов, лежат друг над другом, то нетрудно объяснить быстрые изменения окраски, свойственные многим амфибиям. Зеленый и голубой цвета окраски зависят от сочетания пигментов: зеленый цвет получается при налегании желтых хроматофоров на черные; голубой—от сочетания белого и черного пигментов.
Тепло, солнечный свет, пониженная влажность вызывают обычно посветление окраски; напротив, холод, темнота и высокая влажность вызывают потемнение. Многие земноводные, особенно лягушки, могут более или менее быстро принимать окраску, сходную с окраской окружающих предметов; например, квакши принимают окраску, соответствующую листве, среди которой они находятся. Изменение окраски происходит через посредство нервной системы, на которую оказывают свое влияние как различные условия внешней среды, так и внутреннее состояние организма.
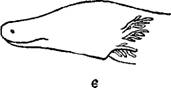
У многих земноводных в эпителии имеются одноклеточные железы, слизистое, вещество которых содействует отделению сбрасываемого во время линьки внешнего слоя эпителия. В кориуме заложено огромное количество многоклеточных желез двоякого рода как по своему .строению, так и по характеру выделений. Одни из них, более крупные, различной величины, расположены исключительно на спинной стороне тела и образуют здесь бородавчатые и другие возвышения кожи; они лежат то неправильно, то продольными лентами, как, например, спинно-боковые валики наших бурых и зеленых лягушек, или группами, как околоушные железы (паротиды) жаб и саламандр. Содержимое выделяющих клеток этих желез зернистое, выделение их млечное, кислое, очень горькое, иногда сильно пахнущее; именно их и называют обыкновенно ядовитыми железами. Железы второго рода меньше, одинаковой величины, рассеяны по всему телу, как по спинной, так и по брюшной стороне его; содержимое этих желез слизистое, выделение жидкое, бесцветное, без особого запаха и со средней или щелочной реакцией. Эти меньшие железы известны большей частью под именем слизеотделительных, хотя выделения их также ядовиты.
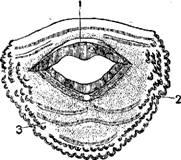
Голова серой жабы (Bufo bufo) с околоушными железами (паротидами).
И те и другие железы одеты снаружи слоем гладких мышечных волокон и открываются на nповерхности кожи короткими, узкими выводными токами яда очень незначительно.
Выделения кожных желез амфибий, пои на слизистые оболочки (например в полости вызывают сильное жжение, а иногда и отравляющее, действие. Известны случаи, когда выдел желез саламандры, попадая на язык собаки, водили к гибели животного; ящерицы, кусая саламандр, умирали в конвульсиях. Введение ядовитых выделений желез земноводных в различных животных чаще всего приводит к гибели последних, даже если количество введенного яда спинных желез обыкновенной саламандры действует на центральную нервную систему и вызывает судороги. Яд гребенчатого тритона замедляет дыхание и вызывает паралич сердца. Яд спиннных желез обыкновенной жабы вызывает столбняк и парализует деятельна сердца. У жаб и саламандр ядовитые выделения соответственно бол численности желез обильнее, чем у других земноводных, и могут еще значительно увеличиваться при раздражении кожи. Содержимое желез мо не только выступать на поверхность кожи, но у некоторых саламандр даже прыскиваться на несколько сантиметров при возбуждении животного.
И все же земноводные не могут считаться ядовитыми животными в по смысле этого слова, так как их ядовитые органы не имеют каких-либо pанящих приспособлений и они не могут напасть на своего врага и причинить ему отравление путем введения яда в кровь. Ядовитые выделения земноводных гут действовать лишь на кожу и слизистые оболочки животных, которые громадном большинстве случаев бывают неповрежденными. Для человека земноводных практически мало опасен, так как никто не станет брать на или саламандру в рот, а если и попытается проделать это, то не повтор своего опыта, испытав однажды сильное жжение языка и слизистых обо чек рта.
Кожный скелет. Вымершие стегоцефалы имели чешуйчатый панцырь, особенно хорошо развитый на брюшной стороне тела, а также мое черепной панцырь из окостенений, образовавшихся в глубоких слоях кожи. Чешуи стегоцефалов имели круглую, овальную или веретенообразную форму и располагались косыми рядами на брюхе, нередко черепицеобразно налегая друг на друга. Незначительные остатки подобного панцыря у современных земноводных имеются у некоторых родов безногих (Apoda); у них в глубоком слое кожи заключены тонкие круглые известковые чешуйки, расположенные, и дранки крыши. Замечательно, что эти чешуйки напоминают по своему строев чешую австралийского чешуйчатника из двоякодышащих рыб. Кожные окостенения в виде щитов на спинной стороне развиваются также у некоторых бесхвостых земноводных; у рогатых лягушек (Ceratophrys) такой костный щит лежит в коже свободно, у седлоносной жабы (Brachycephalus) из семейств» узкоротых лягушек он сливается с позвонками.
Внутренний скелет. Строение скелета земноводных представляет довольно существенные различия в разных группах. У вымерших панцирных земноводных тела позвонков, если они были достаточно окостеневшим и имели вогнутость как спереди, так и сзади (двояковогнутые, или амфицельные, позвонки). Двояковогнутые позвонки свойственны среди современных земноводных безногим и некоторым хвостатым. У других хвостатых и немногих бесхвостых (чесночниц) развивающийся между позвонками хрящ срастается с задней поверхностью позвонков и образует их суставные головки, соответствующие вогнутостям на переднем конце позвонков (передневогнутые, или процельные, позвонки).
Но у большинства бесхвостых межпозвоночный хрящ срастается с передней поверхностью позвонков и образует здесь их суставные головки (задневогнутые, или опистоцельные, позвонки). Число позвонков может доходить до 300 у безногих; значительное число позвонков имеют многие хвостатые (у амфиумы до 100). Напротив, у большинства видов бесхвостых амфибий всего лишь 9 позвонков: 1 шейный, 7 туловищных и 1 крестцовый, позади которого находится палочкообразная копчиковая кость, происшедшая путем слияния значительного числа позвонков. У одной африканской когтеносной лягушки брахиозавра (Brachiosaurus) и чешуи (Hymenochirus) только без позвонков, брюшного панцыря различных других стегозавров, так как сливаются вместе два перцефалов.
Два первых позвонка, а кроме того, крестцовый позвонок срастается и с двумя позвонками, лежащими перед ним, и с копчиковой костью.
У некоторых стегоцефалов и у всех ныне живущих земноводных позвонки, за исключением первого—единственного шейного, имеют боковые поперечные отростки. Длинные тонкие ребра имелись только у одной группы панцирных земноводных, микрозавров, которые приближались также и по присутствию настоящих когтей к пресмыкающимся, но и у них ребра не доходили до грудной кости. Ни одно земноводное не имеет, таким образом, грудной клетки, образованной ребрами. Однако обособленные короткие ребра мы находим у всех саламандрообразных и змеевидных форм, а среди бесхвостых— у круглоязычных (Discoglossidae), притом лишь на самых передних позвонках, а также у личинок безъязычных (Aglossa).
Грудной пояс никогда не бывает плотно соединен с позвоночным столбом.
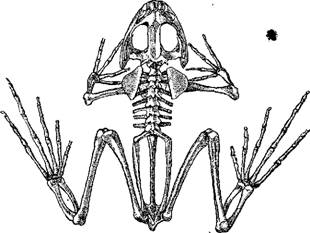
Скелет лягушки (Rana)
Таз прикреплен к одному, редко к двум, трем позвонкам, которые у хвостатых едва отличаются от остальных. У бесхвостых форма поперечных отростков этих позвонков очень характерна для отдельных семейств: они могут быть или круглыми в поперечном сечении или более или менее расширенными и иногда настолько, что представляют собой большую треугольную костную пластинку.
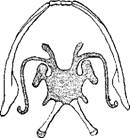
Нижняя челюсть и подъязычная кость лягушки (Rana).
Череп стегоцефалов значительно отличался от черепа других земноводных и был похож на череп кистеперых рыб. Кроющие кости, развившиеся в нижних слоях кожи, слагались в плот состоявшую из многочисленных симметричных кт капсулу, в стенках которой проходили каналы кожных органов чувств; глаз был окружен код из костных пластиночек, так же как у названных уже кистеперых рыб, многих пресмыкающихся и У современных земноводных кроющие кости развиты слабо, и под ними, особенно у бесхвостых, отчасти. сохраняется еще хрящевой первичный череп, между тем как другие части его окостеневают. Череп очень широкий и плоский с очень большими, как прав сквозными глазницами, поэтому при рассматривании сверху челюсти имеют вид дуги, огибающей продолговатую капсулу собственно черепа. Наиболее массивным черепом обладают безногие, у которых он играет существенную роль при рытье. У безногих и у хвостатых земноводных лобные кости обособлены от теменных, между тем как у бесхвостых лобная и теменная кости каждой стороны сливаются между собой, а у безъязычных (Aglossa) обе эти даже срастаются по средней линии. Предлобные кости, которые у многих и у большинства хвостатых земноводных остаются еще обособленными, у бесхвостых или сращены с носовыми костями или вовсе отсутствуют. У последних еще отличимы нёбные кости, которые у хвостатых земноводных срослись с сошниковыми костями или тоже могут отсутствовать. Из двух боковых затылочных костей каждая образует по суставному бугорку мыщелку для соединения с первым шейным позвонком.
Личинки всех земноводных и взрослые особи тех хвостатых, которые в течение всей жизни дышат жабрами, имеют с каждой стороны по 4 жаберные дуги; европейский и американский протей лишь по 3, исполинская саламандре; только по 2, а остальные хвостатые во взрослом состоянии лишь у безногих жаберных скелет по окончании метаморфоза сохраняется: в виде 3—4 изогнутых перекладин; у бесхвостых земноводных он образует широкую хрящевую пластинку, между тем как жаберные дуги совершенно исчезают; у лягушек от этой хрящевой пластинки отходят с каждой стороны длинный рог и 3 отростка, а назад— 2 расходящиеся продолговатые кости, охватывающие гортань.
Конечности пятипалого типа составляют одну из наиболее характерных черт наземных позвоночных, в том числе и амфибий.
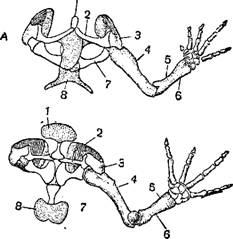
Передние конечности слагаются 8 из простой плечевой кости, двух (у бесхвостых земноводных сросшихся между собой) костей предплечья, часто (у многих хвостатых и стегоцефалов) остающегося хрящевым, запястья, состоящего нормально из 6—8 косточек и большей частью. Плечевой пояс дискоязычной лягушки из 4 пальцев (редко 3, лишь иногда coglossus pictus)— зеленой лягушки (Rana да у амфиумы 2); у форм с меньшим количеством пальцев -esculenta) число косточек запястья сокращается до 2—3.
Сходное строение имеет и задние конечности, в которых у бесхвостых сращены обе берцовые кости. Пяточных костей у хвостатых земноводных 9, но у лягушек меньше, и обе кости ряда, примыкающего к голени, сильно удлинены. Обычно задние конечности имеют 5 пальцев, но у некоторых лягушек и тритонов 4, реже 3 (амфиума) и даже 2 (амфиума, протей).
Пояс передних конечностей, если только последние не отсутствуют совершенно, состоит из трех основных частей: лопатки, переднекоракоидной и заднекоракоидной кости, или коракоида собственно. Лопатка в части, прилежащей к плечу, всегда окостеневает, между тем как вдающаяся в спинные мускулы концевая половина ее остается хотя бы частично хрящевой; коракоидные кости у хвостатых амфибий тоже хрящевые. Коракоидные кости правой и левой сторон могут частично налегать друг на друга и быть взаимно и в известной мере подвижно связанными посредством мускулов, как у тритонов и низших лягушек, которые вследствие этого называются подвижногрудыми; или кости грудного пояса обеих сторон плотно соединены между собой по средней линии хрящом, как у настоящих лягушек, которых поэтому и относят к группе неподвижногрудых.
Переднекоракоидная кость, обычно развитая слабее коракоида, у некоторых узкоротых лягушек (Engystomatidae) совершенно отсутствует; у большинства лягушек на переднекоракоидную кость налегает ключица. Грудная кость, или грудина, в виде маленькой хрящевой пластинки у хвостатых земноводных вдается по средних линии между коракоидными костями; у подвижногрудых лягушек она имеет то же положение, но крупнее; у неподвижногрудых лягушек она лежит позади примыкающих друг к другу коракоидов; часто грудина состоит из костной рукоятки и хрящевой пластинки (мечевидный отросток), вдающейся между брюшными мускулами. Спереди по средней линии к грудному поясу прилегает сходное образование, так называемая предгрудина, или целиком хрящевая или состоящая также из костной рукоятки и хрящевой вершинной пластинки.
Тазовый пояс, прикрепленный к поперечным отросткам крестцового позвонка, слагается из подвздошной, лонной и седалищной костей. Лонная кость остается в течение всей жизни хрящевой. Подвздошные кости бесхвостых земноводных имеют саблеобразную форму; они образуют на заднем конце в соединении с лонными и седалищными костями вертикальную пластинку, с каждой стороны которой находится суставная ямка для бедренной кости.
Мускулатура земноводных бедна сухожилиями; жировая ткань в мышцах также почти совершенно отсутствует. У личинок, а также у тех хвостатых земноводных, которые постоянно дышат жабрами, мускулы туловища и хвоста, как у рыб, разделены на правильные сегменты (миомеры) соответственно числу позвонков и нервов, отходящих от спинного мозга. Менее ясно это расчленение у безногих земноводных и безжаберных хвостатых, а у бесхвостых амфибий оно вообще существует лишь в виде следов.
Нервная система земноводных имеет еще сравнительно невысокое развитие и во многом сходна с нервной системой двоякодышащих рыб.
Головной мозг вытянут в длину в одной плоскости; передний и средний мозг лишь слегка прикрывают спереди и сзади промежуточный. Полушария переднего мозга удлинены и переходят спереди в обонятельные доли, которые сращены по средней линии у бесхвостых, но разделены у хвостатых. Настоящей коры в переднем мозгу еще нет, но стенки его построены сложнее, чем у рыб, и отдельные их участки имеют различное строение. Боковые стенки промежуточного мозга образуют утолщения, так называемые зрительные бугры. Сверху с промежуточным мозгом связана надмозговая железка (эпифиз), которая, судя по величине теменного отверстия в черепе стегоцефалов, была и сильно развита у вымерших земноводных в виде так называемой теменной стороны.
Глаза; через теменное отверстие к этому глазу подходил нерв от промежуточного мозга; у современных фибий рудименты этого органа чувств имеются у хвостых. Снизу от промежуточного мозга отходит ронковидное выпячивание, к которому прилежит мозговой придаток, или гипофиз, играющий существенную роль как железа внутренней секреции. Имеет хорошо развитые зрительные доли в округлых возвышений. В зрительных долях концентруются важнейшие рефлексы: движения конечное глазных мышц, рефлексы осязания и изменения окрасим, а возможно также и дыхательные рефлексы. Мозжечок имеет вид узкой поперечной пластинки, как у двоякодышащих рыб. Продолговатый мозг представляет посредственное продолжение спинного мозга и имеет сверху ромбовидную ямку.
Спинной мозг значительно превышает головное мозг по своей массе. У хвостатых земноводных он простирается до конца хвоста и дает многочисленные нерве в обе стороны; спинной мозг лягушек очень короток, мало продолжается за крестцовым позвонком и дает лишь 10 пар спинных нервов. В связи с развитие парных конечностей в спинном мозгу амфибий имеются плечевое и поясничное вздутия.
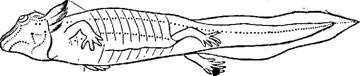
Органы чувств. Органы чувств боковой линии, воспринимающие, вероятно, колебания в давлении воды, имеются у современных земноводных только в личиночном состоянии, а также у постоянно живущих в воде представителей хвостатых и у немногих бесхвостых; у стегоцефалов они были хорошо развиты и по расположению были сходны с органами боковой линии кистеперых рыб.
Органами осязания служат свободные нервные окончания в эпидермисе и специальные осязательные бугорки. С помощью особых нервных окончаний, заложенных в коже, амфибии могут воспринимать колебания температуры окружающей среды и различные химические раздражения. В ротовой полости имеются вкусовые почки.
Органы обоняния представлены двумя разделенными перегородкой полостями, которые открываются спереди ноздрями на конце морды, а сзади сообщаются с ротовой полостью через отверстия (хоаны) на нёбном своде. Подобное строение органов обоняния среди рыб наблюдается' только у двоякодышащих. У многих земноводных отверстия обонятельных полостей могут замыкаться клапанообразными перепонками. У личинок бесхвостых и саламандр хоаны закрыты слизистыми оболочками.
Личинка обыкновенной саламандры (Salamandra salamandra). Точками показаны органы боковой линии.
Расположение органов боковой линии (обозначены черточками) у гладкой шпорцевой лягушки (Xenopus laevis).
Органы слуха у безногих и хвостатых амфибий образованы лишь внутренним ухом, или лабиринтом, который состоит из трех полукружных каналов и мешка, наполненного мелкими кристаллами и имеющего яйцевидное отверстие, закрытое у различных земноводных по-разному. У большинства бесхвостых земноводных имеется также среднее ухо с барабанной перепонкой и слуховой косточкой (столбик). Барабанная полость короткой евстахиевой трубой соединяется с глоткой.
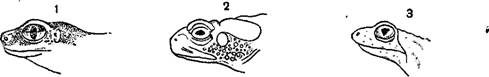
Органы зрения у разных представителей земноводных развиты весьма различно. Лучше всего развиты глаза у бесхвостых: они большей частью велики, очень подвижны, обыкновенно снабжены двумя веками, из которых нижнее больше, тоньше и прозрачнее; в переднем углу глаза имеется, как правило, мигательная перепонка в виде маленькой, обычно неподвижной складки кожи. При помощи особых мускулов глаза могут выпячиваться наружу и втягиваться в глубину орбит. Радужная оболочка у многих земноводных ярко окрашена. Отверстие зрачка в разных группах различно по своей форме, что имеет значение для систематики; у большей части земноводных зрачок горизонтальный, у некоторых ночных форм (чесночницы)—вертикально-эллиптический и днем имеет вид узкой щели, у жерлянок—треугольный. Железы для смачивания глаза имеются у всех амфибий, но настоящих слезных желез они не имеют. Веки развиваются во время метаморфоза; личинки всех земноводных, а также взрослые водные хвостатые век не имеют. У некоторых земноводных глава сильно недоразвиты и скрыты под непрозрачной кожей, как, например, у протея и некоторых безлегочных саламандр (Typhlomolge, Typhlotriton).
Степень развития глаза и форма зрачка у различных земноводных.
1—филломедуза (Phyllomedusa), зрачок вертикально-эллиптический; 2—жаба (Bufo), зрачок горизонтально- эллиптический; 3—жерлянка (Bombina), зрачок треугольный; 4—шпорцевая лягушка (Xenopus), глава малы, но еще имеют веки; 5—червяга (Jchthyophis), глаза малы, просвечивают через кожу; в—протей (Proteus), глаза недоразвиты и совершенно скрыты под кожей.
Органы пищеварения. Разрез рта у большинства а: очень широкий, лишь у немногих бесхвостых ротовое отверстие узкое. расположение и степень развития зубов у разных земноводных весьма разнобразны и имеют значение для систематических различий. Зубы могут вое кости, соприкасающиеся с ротовой полостью.
У хвостатых и безногих земноводных почти всегда (за исключением амфиум) имеются зубы на челюстях; у бесхвостых нижнечелюстные зубы встречаются редко, а в некоторых случаях зубы на челюстях совершенно отсутствуют (жабы, пипа). У большинства земноводных зубы мелкие, полые, конические и служат всегда лишь для удержания пищи и для проталкивания ее в глотку. Они никогда не сидят в луночках, как у млекопитающих и крокодилов, срастаются с челюстными костями; замена их при утрате совершается в течение всей жизни. У личинок бесхвостых челюсти одеты роговой оболочкой образующей подобие клюва.
Язык отсутствует лишь у одного подотряда бесхвостых (Gloss), вообще хорошо развит, часто очень широк, более или менее подвижен и совершенно заполняет пространство между обеими ветвями нижней челюсти. У лягут язык прикреплен передним концом и может поэтому выбрасываться, оборачиваясь нижней стороной вверх. У некоторых хвостатых и бесхвостых, питающихся термитами, он прирос ко дну ротовой полости; у одного рода жаб (Шик nophrynus) язык прикреплен сзади, впереди же свободен и может выдвигать». Наконец, у некоторых земноводных, главным образом у североамериканских наземных саламандр (например Spelerpes), грибообразный язык может далек»; выбрасываться на мускулистом стебельке, как у хамелеонов. У амфибий» живущих постоянно в воде, язык мал, слабо мускулист и мало подвижен, как у рыб.
Пищевод обычно короток и широк, но у безногих земноводных он длинен и узок. Желудок простой, толстостенный, продольно-складчатый, расположенный у хвостатых по длине тела, а у бесхвостых более поперечно. Кишечник разграничен на два отдела: тонкая (двенадцатиперстная) кишка сравнительно коротка и мало извита у взрослых земноводных, но она имеет значительную длину и завита спирально у личинок бесхвостых, которые питаются преимущественно или отчасти растительной пищей; задняя кишка коротка и пузыреобразно расширена; она открывается в клоаку.
Печень обычно большая, разделенная на 2—3 доли. С печенью связан желчный пузырь. Всегда имеется поджелудочная железа. Слюнных желез нет.
Органы дыхания. Основными органами дыхания земноводных служат или легкие или жабры, но, кроме того, в большей или меньшей степени дыхание осуществляется через кожу, а иногда и через покровы ротовой полости и глотки.
Жабрами дышат личинки всех современных амфибий, а также и те взрослые хвостатые, которые всю жизнь проводят в воде. Жаберные дуги имелись у личинок и у некоторых взрослых стегоцефалов. Нормально у земноводных закладываются пять жаберных мешков, но наружу открываются не все, и жаберных щелей никогда не бывает более четырех с каждой стороны, а чаще меньшее число. Внутренние жабры, подобные жабрам рыб, встречаются только у личинок бесхвостых в виде коротких гребневидных выростов покровов на дугах, разделяющих жаберные щели. Как правило, внутренние жабры прикрываются складкой кожи, образующей своего рода жаберную крышку, и притом так. что остается лишь маленькое отверстие, через которое может выходить из жаберной полости вода, поступающая через рот. Таких отверстий может быть или по одному с каждой стороны тела, как у безъязычных бесхвостых (Gloss), или одно непарное на средней линии тела, как у круглоязычных, например у жерлянок (Bombina), или, наконец, .одно несимметричное, лежащее на левой стороне, как у большинства остальных бесхвостых.
Личинки хвостатых амфибий имеют наружные жабры, сходные по своему строению с наружными жабрами личинок двоякодышащих (Dipnoi) и кистеперых рыб (Polypterus). Жабры возникают в виде маленьких бугорков, по бокам головы быстро удлиняющихся и разветвляющихся; во вполне развитом виде они направлены назад и несколько вверх, имеют мускулистый стебель, который может двигаться вверх и вниз, и два ряда жаберных листочков, которые свешиваются вниз, так что верхний край остается свободным. У личинок хвостатых новодных наружные жабры особенно криветвисты; они сидят у верхнего края трехвых жаберных щелей, сохраняются до к превращения и потом медленно атрофиру.
Голова сирена (Siren lacertina) с тремя парами разветвленных жабер.
Личинки бесхвостых сначала также имеют наружные жабры, которые, однако, очень уступают место развивающимся внутренним рам, уже описанным выше.
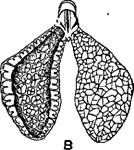
Легкие земноводных очень различны по с форме и строению. У бесхвостых они коротки , приблизительно яйцевидны, пересечены в густой сетью перекладин и перегородок, значительно увеличивает их внутреннюю поверхность, пригодную для дыхания. Наиболее внутреннюю поверхность имеют легкие бесхвостых, ведущих преимущественно наземный образ жизни, например сильно ячеистые легкие наоборот. Кончики больших мешкообразных легких у чесночниц загнуты вперед. У земных саламандр легкие более удлинены, но в остальном такого же строения. У водных хвостатых, дышащих легкими, в последних слабее развита о перегородок. Хвостатые, постоянно живущие в воде и дышащие жабрами; имеют сильно вытянутые гладкостенные легкие, в большой степени утратившие значение органа дыхания и играющие роль гидростатического аппарат, подобно плавательному пузырю рыб. У вытянутых в длину хвостатых и у безногих земноводных правое легкое длиннее левого.
Все представители подсемейства безлегочных саламандр (Plethodontinaef утратили легкие и дышат исключительно посредством очень богатой сосудам ротовой и глоточной полости и кожи. Вентиляция ротовой полости осуществляется путем быстрого опускания и поднимания дна рта; число таких колебаний доходит до 120—170 в минуту.
У большинства земноводных легкие соединяются между собой впереди и открываются непосредственно в глотку продольной щелью, поддерживаемой по сторонам узенькими хрящами; мускулы, прикрепляющиеся к этим хрящам, могут расширять и суживать гортанную щель. Некоторые хвостатые и безногие амфибии имеют довольно длинное дыхательное горло, стенка которого снабжена хрящевыми кольцами. Многие земноводные имеют голосовые связки. У самцов большинства бесхвостых на дне или по углам рта имеются голосовые мешки (резонаторы), раздувающиеся при квакании и способствующие значительному усилению голоса. Личинки всех земноводных совершенно лишены голоса.
В связи с отсутствием грудной клетки легочное дыхание земноводных (вентиляция легких) происходит чрезвычайно своеобразно. Животное втягивает воздух в ротовую полость через ноздри, опуская дно рта с помощью межчелюстной мышцы. Затем ноздри замыкаются клапанами, межчелюстная мышца.
Легкие чесночницы .(Pelobates iuscus)—А, с загнутыми поднимается кверху, и вперед кончиками, и легкие зеленой лягушки (Rana escu дух нагнетается через горло)— В, одно из них в разрезе.
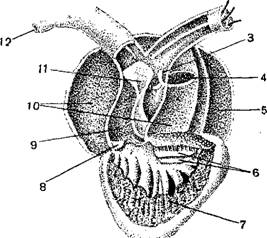
Сердце лягушки (Rana) с брюшной стороны в разрезе.
Обработанные щели в легкие пассивно растягиваются. Из легких воздух выталкивается обратно благодаря спаданию их эластичных стенок.
В области гортани находится щитовидная железа в виде пары очень маленьких телец. Будучи железой внутренней секреции, она играет очень большую роль в регуляции обмена веществ, а также обусловливает наступление метаморфоза у личинок.
Органы кровообращения земноводных обладают рядом особенностей, связанных с переходом от жаберного к легочному дыханию. Характерны еще симметричное расположение артериальных дуг, развитие легочных симметричных артерий и разделение сердца на артериальную и венозную половины (разделение это еще неполное):
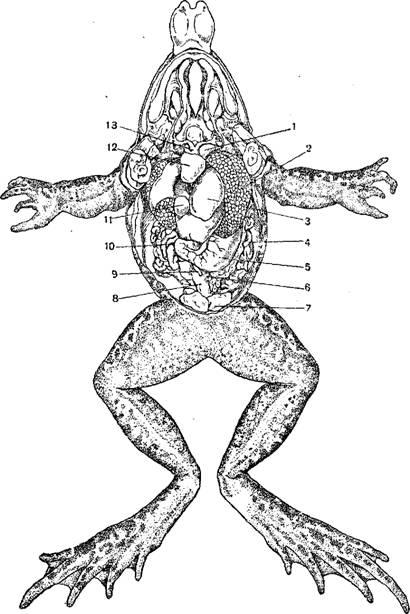
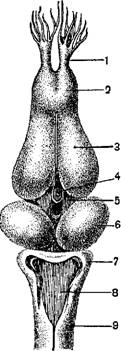
1—«сонная железа»; 2—подъязычная артерия; 3—клапаны; 4—отверстие из венозной пазухи в предсердие; 5— левое предсердие; 6—клапаны между предсердиями и желудочком; 7—желудочек; 8—клапаны; 9—артериальный конус; 10 — правое предсердие; 11—продольная перегородка артериального конуса; 12 — легочнокошная артерия; 13—аорта; 14—сонная артерия.
Сердце расположено в переднем отделе туловища и защищено снизу грудным поясом конечностей; у безногих амфибий оно значительно смещено назад. Сердце заключено в тонкую околосердечную сумку. Оно состоит из толстостенного желудочка, от которого отходит вперед артериальный конус, далее из двух предсердий, лишь у бесхвостых вполне отделенных друг от друга перегородкой, и венозного синуса, принимающего крупнейшие венозные сосуды. В полости желудочка, от его стенок, отходят длинные мускульные тяжи, которые прикрепляются к свободным краям двух клапанов, прикрывающих отверстие между предсердиями и желудочком. Артериальный конус у основания и на конце имеет поперечные клапаны, а внутри разгорожен спиральной перегородкой на верхний—легочный отдел и нижний—артериальный.
Артериальный конус обладает сократимыми стенками и ответвляет вперед не более четырех артерий с каждой стороны, называемых также дугами аорты. У личинок хвостатых в период наличия наружных жабер от артериального конуса отходят четыре пары приносящих жаберных артерий (соответствующие четырем задним из шести артериальных дуг, имеющихся у рыб). Из них первые три идут к жабрам, где распадаются на капилляры, а отсюда снова собираются в отводящие жаберные артерии; последние, направляясь назад, сливаются в спинную аорту. Четвертая пара артериальных дуг прямо переходит в спинную аорту, не распадаясь на капилляры, так как соответственных жабер нет; от этой пары вдут ветви к легким (одна из стадий развития самостоятельных легочных артерий). У личинок саламандр только первая артериальная дуга распадается на капилляры. У хвостатых земноводных, дышащих жабрами в течение всей жизни, первая жаберная артерия обособляется от второй и дает начало сонным артериям, вторая и третья сливаются в аорту, а четвертая в еще большей степени становится легочной артерией. У бесхвостых амфибий третья пара дуг исчезает, вторая пара образует дуги аорты, а четвертая, совершенно теряя связь.
С аортой, в полной мере становится легочной артерией. От первой жаберной артерии отходят вперед к головному мозгу внутренняя сонная артерия языку—подъязычная артерия. У саламандр и бесхвостых при основании артерий находится капиллярное сплетение сосудов. Каждая дуга аорты ответвляет подключичную артерию, идущую в переднюю конечность. От левой дуги аорты отходит чревная артерия, несу кровь к внутренностям. У бесхвостых от каждой легочной артерии отходит крупная кожная артерия, ветвящаяся в коже.
Венозная система земноводных имеет некоторое сходство с венозной системой двоякодышащих рыб. Венозная кровь из головы, от передних конечностей и от кожи собирается с каждой стороны в яремные и подключичные вены, которые сливаются в парные передние полые вены, впадающие в венозный синус. Кровь из задних конечностей поступает в подвздошную вену, разделяющуюся далее на две ветви. Из одной ветви кровь поступает в почки и здесь, разбиваясь на капилляры, принимает участие в образовании воротной системы почек. Капилляры воротной системы сливаются снова и образуют заднюю полую вену, выносящую кровь из почек и впадающую в венозный синус. Вторая ветвь подвздошной вены, сливаясь с соответствующей веной другой стороны, образует брюшную вену. Последняя направляется к печени и вместе с веной, подходящей от внутренних органов, образует воротную систему печени. Из печени кровь выносится двумя печеночными венами, впадающими в заднюю полую вену. Наконец, парные легочные вены, несущие артериальную кровь из легши, соединяются в общую легочную вену, впадающую непосредственно в левое предсердие.
Через «полые вены—две передние и одну заднюю—со всех концов тела собирается венозная кровь и вливается в венозную пазуху. Сокращение стенок пазухи проталкивает кровь в правое предсердие. В то же время в левое предсердие по легочной вене из легких поступает артериальная кровь. При одновременном сокращении обоих предсердий и венозная и артериальная кровь попадает в полость желудочка. При начинающемся расширении (диастоле) предсердий и сокращении (систоле) желудочка отверстия между ними замыкаются клапанами. Венозная кровь попадает в правую половину желудочка, артериальная—в левую. В плоскости их соприкосновения происходит лишь частичное смешение обоих потоков крови, так как этому препятствуют ряд мышечных перекладин на внутренних стенках желудочка и краткость самой диастолы желудочка. В первой фазе сокращения желудочка кровь устремляется в открывающийся к этому моменту артериальный конус. Последний находится на правой стороне основания желудочка, и вполне естественно, что первая порция крови, попадающая в него, будет венозной, так как она заполняет именно правую половину желудочка. Из артериального конуса кровь поступает в легочные артерии, отверстия которых в это время широко открыты, и заполняет их. Во второй фазе систолы желудочка происходят сокращение стенок артериального конуса и перемещение его спирального клапана, который закрывает легочный отдел конуса и оставляет открытым нижний артериальный. Вторая порция крови, смешанная по своему составу, поступает в дуги аорты, но не проникает в сонные артерии, встречая здесь более высокое давление, создаваемое капиллярами «сонных желез». В третьей фазе сокращения желудочка, когда гонится из его полости чисто артериальная кровь, в дугах aoрты сопротивление повышено предшествующим наполнением, и поток крови, наконец, преодолевая давление в сонных артериях, заполняет их. Таким образом, последняя порция крови, чисто артериальная, попадает в сонные артерии, омывающие мозг.
Красные кровяные клетки—эритроциты—эллиптической формы и сравнительно очень велики. Селезенка, имеющая вид небольшого круглого тела красного цвета, располагается на брюшине вблизи начала прямой кишки.
У земноводных сильно развита лимфатическая система, состоящая из щелевидных пространств. Бесхвостые имеют под кожей большие отделенные друг от друга перегородками лимфатические полости. Поэтому кожа прикрепляется к находящейся под ней мускулатуре лишь в тех местах, где находятся эти перегородки. Лимфатические сосуды открываются в большие венозные стволы и расширены в этом месте в так называемые лимфатические сердца, пульсирующие самостоятельно. Большинство бесхвостых земноводных имеет две пары таких лимфатических сердец: одну в плечевой области, другую поблизости клоачного отверстия на спинной стороне.
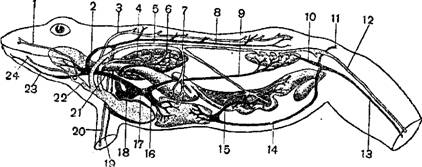
Органы выделения. Почки, расположенные по обе стороны позвоночника, у хвостатых земноводных сильно удлинены и состоят из двух отделов: из переднего, узкого, связанного у самцов с половыми железами, и заднего, более широкого, который является собственно почкой. У бесхвостых почки не имеют такого подразделения и в виде эллиптических тел расположены примерно по середине туловища. Мочеточники открываются в клоаку. Объемистый, тонкостенный, часто двурогий мочевой пузырь сообщается с брюшной стенкой клоаки. Наполнение его мочой Происходит через клоаку; через клоаку же происходит Лимфатические пространства под кожей лягушки (А—со спинной стороны, В—с брюшной стороны и его опорожнение)
Темными полосами обозначены перегородки между отдельными лимфатическими мешками являющиеся важными железами внутренней секреции.
Мочеполовая система самца лягушки (Discoglossus pictus) — А и зеленой лягушки (Rana esculenta) — В.
Органы размножения у земноводных в значительной степени связаны с органами выделения, поэтому обычно обе системы рассматриваются как мочеполовая система. Всегда парные половые железы прикрепляются к спинной полости тела на мезентерии.
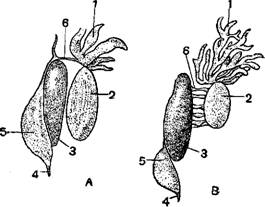
Мужские половые железы-семенники—у хвостатых амфибий вытянуты в длину, у бесхвостых имеют овальную форму. Поло-продукты выводятся из семенников через тонкие выводные канальцы, которые так или иначе впадают в выводные каналы (мочеточники), называющиеся поэтому нередко мочесеменными каналами. У хвостатых выводные каналы впадают в мочевые канальцы переднего узкого отдела почек; у большинства бесхвостых они также проходят через почку, но у некоторых круглоязычных (Discoglossidae) открываются непосредственно в мочеточник. У бесхвостых амфибий в период размножения мочеточники перед впадением в клоаку образуют расширения—семенные пузырьки, служащие резервуарами для семени. У безногих земноводных клоака может выворачиваться и служит органом совокупления.
Женские половые железы—яичники—вполне отделены от остальных органов полового аппарата. У хвостатых они имеют вид удлиненного мешка с единственным отверстием, через которое яйца выводятся в полость тела. У бесхвостых яичники гроздевидны, и каждое яйцо созревает в отдельной капсуле разрывающейся при выходе яйца. Яйцеводы очень длинны (у бесхвостый; сильно извиты) и открываются в полость тела широкими воронками у основании легких; их конечные отделы часто представляют расширение, похожее на матку, в котором у живородящих саламандр и некоторых лягушек действительно происходит развитие зародышей. Яйцеводы правой и левой сторон открываются в клоаку самостоятельными отверстиями. Яйца из полости тела попадают в воронки яйцеводов и, проходя через последние, одеваются оболочками, сильна разбухающими в воде. У самок саламандр имеются семенные мешки (семе-приемники) в стенке клоаки, в которых семя может оставаться живым долгое время после спаривания.
У всех земноводных в связи с половыми железами находятся лентовидные или лопастные жировые тела, обычно имеющие яркую желтую окраску. Наибольшего развития они достигают осенью перед зимовкой и сильно уменьшаются в величине после периода размножения. Жировые тела служат для питания половых желез и развивающихся в них половых клеток.
Все земноводные раздельнополы. Однако у самцов обычно имеются недоразвитые яйцеводы. У жаб (Bufo) в связи с передним отделом половых желез семенника (или яичника) имеется особая железа, получившая название биддерова органа. Если у самца жабы произвести операцию удаления семенников, при сохранении биддеровых органов можно добиться превращения его в самку. При этом биддеровы органы развиваются в яичники, а недоразвитые яйцеводы получают полное развитие.
Стадии развития личинки лягушки (Rana). А — личинка перед вылуплением; В — личинка, еще не утратившая наружных жабер; С—головастике хорошо развитыми задними конечностями; через кожу просвечивают передние конечности, сердце и кишечник.
Яйца земноводных довольно богаты желтком. В яйце лягушки можно различить две неравные половины. Одна из них, несколько меньшая и пигментированная, заключает главную массу протоплазмы; эту половину называют анимальным полюсом. Другая, большая и светлоокрашенная половина яйца выполнена желтком и носит название вегетативного полюса. После оплодотворения, которое у огромного большинства земноводных происходит во внешней среде, начинается развитие яйца. Дроблению на все возрастающее число клеточек (бластомеров) подвергается без остатка вся масса яйца (так называемое полное дробление), но образующиеся бластомеры неодинаковой величины, и те из них, которые находятся на вегетативном полюсе, значительно крупнее бластомеров анимального полюса (неравномерное дробление).
У некоторых хвостатых земноводных яйца содержат особенно много желтка, и дробление их приближается к частичному дроблению. В общем дробление яйца у земноводных сходно с дроблением у двоякодышащих рыб.
Первая борозда дробления проходит вертикально и делит яйцо на два полушария. Плоскость этого первого деления является вместе с тем и плоскостью симметрии будущего зародыша. Вторая борозда тоже вертикальная и перпендикулярная к первой. Третья борозда горизонтальна и лежит ближе к анимальному полюсу. Затем яйцо продолжает дробиться вертикальными и горизонтальными бороздами и притом быстрее на анимальном полюсе, чем на вегетативном. Мы опустим дальнейшее описание ранних стадий развития и сразу перейдем к тем стадиям, когда в яйцевых оболочках можно уже различить сформировавшийся зародыш.
Зародыш образуется через несколько дней после оплодотворения яйца, а у дискоязычных (Discoglossidae) уже в течение 30 часов. Сначала он довольно короткий, полулунно-изогнутый, с толстой головой и очень коротким хвостом; позднее хвост удлиняется, сплющивается с боков и приобретает кожную складку, окаймляющую его как сверху, так и снизу. На более поздних стадиях развития можно обнаружить мускулы хвоста, расположенные такими же зигзагообразными пластинками, как у рыб. Ротовое отверстие намечается первоначально в виде поперечного или ромбоидального углубления на переднем конце зародыша. Позади рта на брюшной стороне находится очень различно развитый у разных видов орган прикрепления, имеющий у бесхвостых земноводных подковообразную форму; не являясь присоской, этот орган выделяет липкую жидкость, с помощью которой зародыш удерживается на студенистой оболочке яйца, а позднее на водяных растениях. О развитии жабер было уже сказано выше.
Личинки бесхвостых земноводных, прежде чем достигнут окончательного вида взрослых животных, подвергаются гораздо более значительному превращению, чем личинки хвостатых; у них не только жабры, но в связи с переменой танин и рот и кишечник испытывают глубокие изменения. Как уже было, первоначальные наружные перистые жабры скоро заменяются так называемыми внутренними, которые отделяются от окружающей среды выращеной кожной складкой. В то же время развивается кишка, быстро удлиняющаяся и свертывающаяся в виде спирали; она просвечивает сквозь кожу и латуру на брюшной стороне туловища.
Рот получает роговые челюсти, а «губы»—мелкие роговые зубчики, и прилипания исчезает. Затем образуются зачатки задних ног, которые постепенно увеличиваются в длину до полного развития молодой лягушки. Одновременно образуются и передние конечности, но они остаются до полного развития скрытыми в мешке жаберной полости, а прорывают кожу или, если на левой стороне тела имеется жаберное отверстие, то через него часто высовывается наружу соответственная передняя нога. Далее постепенно укорачивается хвост, а кожная кайма его исчезает. Жабные дуги утрачиваются. Легкие, которые чала лишь отчасти служили для дыхания, отчасти для регулирования равновесия в воде, теперь переходят исключительно к дыхательной функции. Роговое вооружение челюстей сбрасывается кусками. Лежавшие сначала под прозрачной надкожицей глаза становятся свободными и получают веки. Разрез рта становится широким; кишка укорачивается. Наконец молодое бесхвостое земноводное оставляет воду, по большей части имея остаток хвоста.
Развитие проходит не всегда одинаково в различных отрядах и ceмействах. Некоторые земноводные или более или менее значительную часть даже все свое личиночное развитие проходят в яйце и притом вне воды. Сравнительно немногие амфибии живородящи. Замечательно, что во всех этих случаях особенно различно строение личиночных органов дыхания.
Обыкновенная саламандра рождает детенышей с тремя парами жабер; личинки заканчивают свое превращение в воде. Жабры личинок альпийской саламандры, развивающихся в теле матери, чрезвычайно велики тонки, уплощены, по краю бахромчаты, богаты кровеносными сосудами лишены пигмента, но в остальном не отличаются существенно от жабер личинок, живущих в воде. В других случаях жабры, напротив, кожисты и пронизаны многочисленными разветвлениями кровеносных сосудов. У сумчатых квакш (Nototrema) она колоколовидны с двумя длинными стебельками, идущими от второй и третьей жаберных дуг; в каждом стебельке проходит по одной артерии и одной вене, ветвящихся в колоколе. Всего лишь одну пару кожистых листовидных жабер имеют личинки некоторых безногих (Typhlonectes). У листовых лягушек (Hylodes) и пипы вообще нет жабер в личиночном состоянии, и органом дыхания в этих случаях служит богатый кровеносными сосудами хвост. Головастик водяной лягушки с Соломоновых островов (Rana opisthodon) дышит при помощи девяти пар кожистых складок по бокам брюха.
Для некоторых бесхвостых земноводных (озерная лягушка, чесночница, жаба-повитуха и некоторые другие) известны случаи, когда их головастики мандры (Salamandra atrai не заканчивают своего превращения в нормальные сроки, а остаются еще год или более в личиночном состоянии, достигая при этом очень значительной еще до рождения.
Молодая сумчатая квакша (Nototre- ma cornutum) с колоколовидными жабрами, извлеченная из выводковой сумки.
Личинки некоторых хвостатых земноводных не только могут достигать величины взрослых особей, но оказываются также способными к размножению, не проделав метаморфоза. Это явление сохранения личиночных признаков в половозрелом состоянии получило название неотении. Широко известным примером неотении является аксолотль—личинка амблистомы, но, кроме того, могут быть названы некоторые виды тритонов (Triturus), у которых также наблюдалось размножение, когда все характерные признаки личинки имелись еще в полной степени. Весьма вероятно, что сохраняющие в течение всей жизни жабры амфиумовые (Amphiumidae) и некоторые безлегочные саламандры (Plethodontinae), подобно аксолотлю, являются неотеническими личинками, наземные формы которых нам остаются неизвестными в связи с отсутствием у них метаморфоза.
Ряд замечательных исследований, произведенных за последние 25 лет, показал огромную роль щитовидной железы в развитии амфибий, а также в значительной мере выяснил причины неотении.
Гудернэч своими опытами в 1912 г. установил, что если подкармливать головастиков кусочками щитовидной железы, взятой от любого животного, то можно получить ускоренное превращение головастиков в лягушек. При этом часто получаются лягушата карликовых размеров. Тех же результатов можно достигнуть, если к воде, в которой живут головастики, прибавить вытяжку из щитовидной железы, заключающую в себе ее выделения (гормон), так называемый тироксин. Позднее было установлено, что головастики, у которых была удалена или разрушена щитовидная железа, продолжают расти и достигают гигантских размеров и даже приступают к размножению, но утрачивают способность к метаморфозу.
Личинка безногой амфибии (Typhlonectes) с одной парой листовидных жабер. Личинка извлечена из яйцевой оболочки в гонце развития.
Опыты с подкармливанием щитовидной железой аксолотлей (Лауфберга, Н. Кольцова, Б. Завадовского и других) дали не менее замечательные результаты. Приблизительно через месяц после начала кормления молодой аксолотль утрачивает жабры и гребень на спине и хвосте, приобретает иное сложение тела и пятнистую окраску и превращается, таким образом, в амблистому, переходящую к наземному образу жизни. Сравнительно недавно гистологическими исследованиями было выяснено недоразвитие щитовидной железы аксолотля.
Как только что приведенными, так и многими другими исследованиями было установлено, что гормон щитовидной железы, тироксин, поступая в личинки, в определенные периоды ее развития обусловливает нас метаморфоза. Недоразвитие щитовидной железы в силу тех или иных еще; выясненных причин нарушает нормальный ход развития амфибий—метаморфоз не наступает, но рост не останавливается, а половые железы развиваются нормально. В результате получаются неотенические личинки.

 Расположение органов боковой линии (обозначены черточками) у гладкой шпорцевой лягушки (Xenopus laevis).
Расположение органов боковой линии (обозначены черточками) у гладкой шпорцевой лягушки (Xenopus laevis).